The New Cells Must Immediately Begin to Divide Again
The jail cell bike culminates in the partition of the cytoplasm by cytokinesis. In a typical prison cell, cytokinesis accompanies every mitosis, although some cells, such as Drosophila embryos (discussed afterwards) and vertebrate osteoclasts (discussed in Chapter 22), undergo mitosis without cytokinesis and become multinucleate. Cytokinesis begins in anaphase and ends in telophase, reaching completion as the adjacent interphase begins.
The first visible change of cytokinesis in an animal cell is the sudden advent of a pucker, or cleavage furrow, on the cell surface. The furrow rapidly deepens and spreads effectually the cell until it completely divides the jail cell in two. In animal cells and many unicellular eucaryotes, the structure that accomplishes cytokinesis is the contractile ring—a dynamic assembly composed of actin filaments, myosin Two filaments, and many structural and regulatory proteins. The ring assembles just below the plasma membrane and contracts to constrict the cell into two (see Figure 18-four). At the aforementioned fourth dimension, new membrane is inserted into the plasma membrane adjacent to the contractile ring by the fusion of intracellular vesicles. This addition of membrane is required to compensate for the increase in surface area that accompanies cytoplasmic division. Thus, cytokinesis can be considered to occur in four stages—initiation, contraction, membrane insertion, and completion.
The fundamental problem for a cell undergoing cytokinesis is to ensure that information technology occurs at the right time and in the correct place. Cytokinesis must non occur too early in Thousand-stage, or information technology volition disrupt the path of the separating chromosomes. It must also occur at the right place to separate the ii segregating sets of chromosomes properly and so that each daughter cell receives a complete ready.
The Microtubules of the Mitotic Spindle Decide the Plane of Animal Cell Division
The mitotic spindle in animal cells not only separates the girl chromosomes, it likewise specifies the location of the contractile ring, and thereby the plane of cell division. The contractile band invariably forms in the plane of the metaphase plate, at right angles to the long axis of the mitotic spindle, thereby ensuring that division occurs betwixt the two sets of separated chromosomes. The part of the spindle that specifies the sectionalisation plane varies depending on the cell type: in some cells, it is the astral microtubules; in others, it is the overlapping antiparallel microtubules in the primal spindle.
The relationship between the spindle microtubules and the placement of the contractile band has been studied by manipulating fertilized eggs of marine invertebrates. Afterwards fertilization, these embryos undergo a serial of rapid cleavage divisions, without intervening periods of growth. In this way, the original egg is progressively divided upwards into smaller and smaller cells. During cytokinesis, the cleavage furrow appears of a sudden on the surface of the cell and deepens rapidly (Figure 18-30). Because the cytoplasm is clear, the spindle tin exist observed in real time through a microscope. If the spindle is tugged into a new position with a fine drinking glass needle in early anaphase, the incipient cleavage furrow disappears, and a new i develops in accordance with the new spindle site.

Effigy eighteen-30
Cleavage in a fertilized frog egg. In these scanning electron micrographs, the cleavage furrow is especially obvious and well defined, as the cell is unusually large. The furrowing of the prison cell membrane is caused past the activity of the contractile ring (more...)
How does the mitotic spindle control the plane of division? Ingenious experiments in large embryonic cells demonstrate that a cleavage furrow forms midway between the asters originating from the two centrosomes, even when the two centrosomes are not connected to each other past a mitotic spindle (Figure eighteen-31). Thus, in these cells, the microtubule asters—not the chromosomes or other parts of the spindle—indicate to the cell cortex to specify where the contractile ring should get together. In other cells, the central spindle, rather than the astral microtubules, is obviously responsible for this specification. In either case, it has been speculated that the overlapping microtubules may provide tracks for motor proteins to deliver contractile ring regulators, and perhaps new membrane, to the appropriate region of the dividing cell. But, in fact, the molecular machinery by which the spindle positions the cleavage furrow remains a mystery.

Figure eighteen-31
An experiment demonstrating the influence of the position of microtubule asters on the subsequent plane of cleavage in a large egg cell. If the mitotic spindle is mechanically pushed to one side of the prison cell with a glass dewdrop, the membrane furrowing is (more...)
In some cells, the site of ring assembly is chosen before mitosis, co-ordinate to a landmark placed in the cortex during a previous prison cell cycle. In budding yeasts, for example, a ring of proteins chosen septins assembles before mitosis, adjacent to a bud scar left on the cell surface as the female parent and girl cells separated in the previous division. The septins are thought to form a scaffold onto which other components of the contractile band, including myosin 2, assemble. Equally we discuss afterward, in found cells, an organized band of microtubules and actin filaments assembles but before mitosis and marks the site where the prison cell wall will assemble and divide the jail cell in two.
Some Cells Reposition Their Spindle to Carve up Asymmetrically
Virtually cells divide symmetrically. In almost animate being cells, for example, the contractile ring forms around the equator of the parent jail cell, so that the two girl cells produced are of equal size and take similar properties. This symmetry results from the placement of the mitotic spindle, which in most cases tends to center itself in the cytoplasm. The centering process depends both on astral microtubules and on motor proteins that either push or pull on the astral microtubules to center the spindle.
At that place are many instances in development, however, when cells split up asymmetrically to produce two cells that differ in size, in the cytoplasmic contents they inherit, or in both. Usually, the two daughter cells are destined to develop forth different pathways. To create daughter cells with unlike fates, the mother jail cell must first segregate some components (called fate determinants) to one side of the prison cell and so position the aeroplane of division then that the appropriate daughter cell inherits these components (Figure xviii-32). To position the plane of sectionalization asymmetrically, the spindle has to be moved in a controlled manner within the dividing prison cell. It seems likely that such spindle movements are directed by changes in local regions of the prison cell cortex and that motor proteins localiazed there pull one of the spindle poles, via its astral microtubules, to the advisable region (Figure 18-33). Some of the proteins required for such asymmetrical divisions have been identified through genetic analyses in C. elegans and Drosophila (discussed in Chapter 21), and some of these seem to take a similar role in vertebrates.

Figure 18-32
An asymmetric cell division segregating cytoplasmic components to but i daughter cell. These light micrographs illustrate the controlled asymmetric segregation of specific cytoplasmic components to one daughter cell during the first division of a fertilized (more...)

Figure 18-33
Spindle rotation. (A) A possible machinery underlying the controlled rotation of a mitotic spindle. The ruddy bar represents a specialized region of cell cortex toward which one spindle pole is pulled by its astral microtubules. (B) Fluorescence micrographs (more...)
Asymmetric sectionalisation is specially important in plant cells. As these cells cannot move after partition, the selection of division planes is crucial for controlling tissue morphology. Nosotros discuss later how the aeroplane of division is adamant in these cells.
Actin and Myosin II in the Contractile Band Generate the Force for Cytokinesis
As the astral microtubules in anaphase become longer lived and less dynamic in response to the loss of M-Cdk activity, the contractile ring begins to gather beneath the plasma membrane. Much of the preparation for cytokinesis, however, happens before in mitosis, before the partition of the cytoplasm actually begins. In interphase cells, actin and myosin filaments are assembled into a cortical network and, in some cells, also into large cytoplasmic bundles chosen stress fibers (discussed in Chapter 16). As cells enter mitosis, these arrays disassemble; much of the actin is reorganized, and myosin II filaments are released. As the chromatids separate in anaphase, myosin II begins to accumulate in the apace assembling contractile ring (Figure 18-34).

Figure eighteen-34
The contractile ring. (A) A cartoon of the cleavage furrow in a dividing cell. (B) An electron micrograph of the ingrowing edge of a cleavage furrow of a dividing animal cell. (C) Fluorescence micrographs of a dividing slime mold amoeba stained for actin (more...)
In many cells, cytokinesis requires the activation of 1 or more members of the polo-like family of protein kinases. These kinases regulate the assembly of both the mitotic spindle and the contractile ring and are therefore thought to assistance coordinate mitosis and cytokinesis, but it is uncertain how they do so. The fully assembled contractile ring contains many proteins in addition to actin and myosin Ii. The overlapping arrays of actin filaments and bipolar myosin II filaments, even so, generate the forcefulness that divides the cytoplasm in 2. They are idea to contract by a mechanism that is biochemically like to that used by shine musculus cells; in both cases, for example, the contraction begins when Ca2+-calmodulin activates myosin light-chain kinase to phosphorylate myosin 2. Once contraction has been stimulated, the ring develops a strength large enough to bend a fine drinking glass needle that is inserted in the path of the constricting ring.
How the contractile band constricts is withal a mystery. It seems not to operate by a elementary "purse-string" mechanism, with actin and myosin II filaments sliding by each other every bit in skeletal muscle (run across Figure 16-71). As the ring constricts, the band maintains the same thickness in cross-section, suggesting that its total volume and the number of filaments it contains decrease steadily. Moreover, unlike in musculus, the actin filaments in the band are highly dynamic, and their organization changes extensively during cytokinesis.
In addition to specifying the site of contractile ring assembly in early anaphase, in many cells, microtubules besides work continuously during anaphase and telophase to stabilize the advancing cleavage furrow. Drugs that depolymerize microtubules, for example, cause the actin filaments in the contractile ring to become less organized. Moreover, if a needle is used to tear microtubules away from the cell cortex, the contractile ring disassembles and the cleavage furrow regresses. It is non known how the microtubules stabilize the ring, although it has been shown that growing microtubules tin activate some members of the Rho family of pocket-sized GTPases, which in turn stimulate actin polymerization (discussed in Chapter 16). One member of this family unit, Rho A, is required for cytokinesis.
The contractile band is finally dispensed with altogether when cleavage ends, as the plasma membrane of the cleavage furrow narrows to form the midbody. The midbody persists every bit a tether between the two daughter cells and contains the remains of the central spindle, which at present consists of the two sets of antiparallel overlap microtubules packed tightly together inside a dense matrix fabric (Effigy 18-35). Remarkably, in some cells, before cytokinesis has been completed, the female parent centriole from one or both daughter cells separates from its daughter centriole (run into Figure 18-5c) and migrates into the midbody, where it lingers for minutes, before returning to its girl jail cell. Only then exercise the two daughter cells divide to consummate cytokinesis. What the centriole might do in the midbody to trigger the final steps of cytokinesis is not known. Later on the daughter cells separate completely, some of the components of the rest midbody often remain on the inside of the plasma membrane of each cell, where they may serve as a marker on the cortex that helps to orient the spindle in the subsequent cell division.

Figure 18-35
The midbody. (A) A scanning electron micrograph of an animate being cell in culture in the process of dividing; the midbody notwithstanding joins the two daughter cells. (B) A conventional electron micrograph of the midbody of a dividing animal cell. Cleavage is most (more...)
Membrane-enclosed Organelles Must Be Distributed to Daughter Cells During Cytokinesis
The process of mitosis ensures that each daughter cell receives a full complement of chromosomes. But when a eucaryotic cell divides, each girl jail cell must too inherit all of the other essential prison cell components, including the membrane-enclosed organelles. Equally discussed in Chapter 12, organelles like mitochondria and chloroplasts cannot assemble spontaneously from their individual components; they can arise only from the growth and partition of the preexisting organelles. Similarly, cells cannot make a new endoplasmic reticulum (ER) unless some function of information technology is already present.
How, and so, are the diverse membrane-enclosed organelles segregated when a cell divides? Organelles such as mitochondria and chloroplasts are commonly present in large plenty numbers to be safely inherited if, on average, their numbers roughly double once each cycle. The ER in interphase cells is continuous with the nuclear membrane and is organized by the microtubule cytoskeleton. Upon entry into M stage, the reorganization of the microtubules releases the ER, which fragments as the nuclear envelope breaks down. The Golgi apparatus probably fragments every bit well, although in some cells it seems to redistribute transiently into the ER, merely to re-emerge at telophase. Some of the organelle fragments associate with the spindle microtubules via motor proteins, thereby hitching a ride into the daughter cells every bit the spindle elongates in anaphase.
Mitosis Can Occur Without Cytokinesis
Although nuclear division is usually followed past cytoplasmic division, in that location are exceptions. Some cells undergo multiple rounds of nuclear sectionalization without intervening cytoplasmic segmentation. In the early Drosophila embryo, for example, the first 13 rounds of nuclear partitioning occur without cytoplasmic division, resulting in the formation of a single large cell containing 6000 nuclei, arranged in a monolayer almost the surface (Effigy 18-36). This system profoundly speeds up early evolution, as the cells exercise not have to accept the fourth dimension to become through all the steps of cytokinesis for each division. After these rapid nuclear divisions, cells are created around each nucleus in ane round of coordinated cytokinesis called cellularization. Contractile rings form at the cell surface, and the plasma membrane extends inward and pinches off to enclose each nucleus.

Figure 18-36
Mitosis without cytokinesis in the Drosophila embryo. (A) The get-go thirteen nuclear divisions occur synchronously and without cytoplasmic division to create a large syncytium. Most of the nuclei so migrate to the cortex, and the plasma membrane extends (more...)
Nuclear division without cytokinesis also occurs in some types of mammalian cells. Osteoclasts, trophoblasts, and some hepatocytes and heart muscle cells, for example, become multinucleated in this way.
The Phragmoplast Guides Cytokinesis in Higher Plants
Near higher-plant cells are enclosed by a semirigid cell wall, and their mechanism of cytokinesis is different from that just described for creature cells. Rather than a contractile ring dividing the cytoplasm from the outside in, the cytoplasm of the institute prison cell is partitioned from the within out past the construction of a new cell wall, chosen the jail cell plate, betwixt the two daughter nuclei (Figure xviii-37). The orientation of the cell plate determines the positions of the 2 daughter cells relative to neighboring cells. It follows that altering the planes of cell sectionalisation, together with enlargement of the cells by expansion or growth, leads to different cell and tissue shapes that aid make up one's mind the form of the establish.

Effigy eighteen-37
Cytokinesis in a plant prison cell in telophase. In this low-cal micrograph, the early cell plate (between the two arrowheads) is forming in a airplane perpendicular to the plane of the page. The microtubules of the spindle are stained with aureate-labeled antibodies (more...)
The mitotic spindle by itself is non sufficient to determine the exact position and orientation of the cell plate. The showtime visible sign that a college-plant cell has become committed to split up in a particular plane is seen in G2, when the cortical array of microtubules disappears in preparation for mitosis. At this time, a circumferential band of microtubules and actin filaments forms a ring around the entire cell only beneath the plasma membrane. Because this cytoskeletal array appears before prophase begins, information technology is called the preprophase band. The band becomes thinner every bit the prison cell progresses to prophase, and it disappears completely before metaphase is reached. Notwithstanding, the sectionalisation airplane has somehow been established: when the new cell plate forms later during cytokinesis, it grows outward to fuse with the parental wall precisely at the zone formerly occupied by the preprophase band. Even if the jail cell contents are displaced past centrifugation after the preprophase band has disappeared, the growing cell plate tends to find its fashion back to the airplane defined by the quondam preprophase band.
The associates of the cell plate begins in belatedly anaphase and is guided past a construction chosen the phragmoplast, which contains the remaining overlap microtubules of the mitotic spindle that interdigitate at their growing plus ends. This region of overlap is similar in structure to the primal spindle in animal cells in late anaphase. Modest vesicles, largely derived from the Golgi apparatus and filled with polysaccharide and glycoproteins required for the synthesis of the new cell-wall matrix, are transported along the microtubules to the equator of the phragmoplast, apparently by the action of microtubule-dependent motor proteins. Here, the vesicles fuse to form a disclike, membrane-enclosed structure called the early prison cell plate (see Effigy 18-9G). The plate expands outward by further vesicle fusion until it reaches the plasma membrane and the original cell wall and divides the jail cell in two. After, cellulose microfibrils are laid downward within the matrix of the prison cell plate to complete the construction of the new cell wall (Figure eighteen-38).

Figure eighteen-38
The special features of cytokinesis in a higher constitute prison cell. The division plane is established before Grand phase by a band of microtubules and actin filaments (the preprophase band) at the cell cortex. At the outset of telophase, after the chromosomes (more...)
The Elaborate Yard Stage of Higher Organisms Evolved Gradually from Procaryotic Fission Mechanisms
Procaryotic cells divide past a process called binary fission. The single, circular Deoxyribonucleic acid molecule replicates and partitioning occurs by the invagination of the plasma membrane and the laying downwardly of new cell wall between the two chromosomes to produce ii separate daughter cells. In E. coli, before the chromosome replicates, the single origin of replication (oriC) is located at i pole of the rod-shaped bacterium. As soon every bit oriC is replicated, i copy of the sequence is immediately translocated to the reverse pole of the cell, after which the residue of the chromosome is replicated. Similar the two spindle-pole asters in an animal cell, the bacterial daughter chromosomes at the cell poles somehow determine the location of the aeroplane of prison cell sectionalization, ensuring that fission takes place at the cell equator, so that each daughter cell inherits i chromosome (Figure 18-39). Although a number of genes and proteins involved have been identified, the mechanisms responsible for the active translocation of oriC and the inhibition of fission everywhere but at the equator remain unknown.
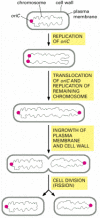
Figure 18-39
Jail cell division in the bacterium East. coli. The single, round chromosome contains an origin of replication chosen oriC. Before division, the chromosome is polarized, and then that oriC is at 1 pole of the bacterium. As shortly as the oriC sequence is copied, (more...)
Binary fission in procaryotes depends on filaments fabricated of the FtsZ protein. FtsZ is a cytoskeletal GTPase that is structurally related to tubulin and assembles into a ring at the equator of the jail cell (Figure xviii-40A, and see Figure 16-17). The FtsZ filaments are essential for the recruitment of all the other prison cell segmentation proteins to the division site. Together, these proteins guide the in growth of the cell wall and membrane, leading to the germination of a septum that divides the cell into two. Bacteria in which the ftsZ factor is inactivated by mutation cannot divide. A FtsZ-based mechanism is also used in the division of chloroplasts in establish cells (Figure eighteen-40B) and mitochondria in protists. In fungi and brute cells, another cocky-assembling GTPase called dynamin (discussed in Affiliate xiii) has apparently taken over the role of FtsZ in mitochondrial division.
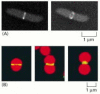
Effigy xviii-40
The FtsZ protein. (A) Fluorescence micrographs showing the location of the FtsZ protein during binary fission in E. coli. The protein assembles into a ring at the center of the cell, where it helps orchestrate cell division. The bacteria here accept been (more...)
With the evolution of the eucaryotes, the genome increased in complexity, and the chromosomes increased in both number and size. For these organisms, a more elaborate mechanism for dividing the chromosomes betwixt girl cells was manifestly required. Clearly, the mitotic apparatus could non take evolved all at once. In many primitive eucaryotes, such every bit the dinoflagellate Cryphthecodinium cohnii, mitosis depends on a membrane-attachment mechanism, in which the chromosomes have to demark to the inner nuclear membrane for segregation. The intermediate status of this large, single-celled alga is reflected in the composition of its chromosomes, which, like those of procaryotes, have relatively little associated protein. The nuclear membrane in C. cohnii remains intact throughout mitosis, and the spindle microtubules remain entirely outside the nucleus. Where these spindle microtubules press on the outside of the nuclear envelope, the envelope becomes indented in a serial of parallel channels (Figure 18-41). The chromosomes go attached to the inner membrane of the nuclear envelope contrary these channels, and chromosome segregation occurs on the within of this channeled nuclear membrane. Thus, the extranuclear "spindle" is used to gild the nuclear membrane and thereby define the plane of division. Kinetochores in this species seem to be integrated into the nuclear membrane and may therefore take evolved from some membrane component.
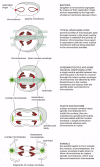
Figure 18-41
The utilize of different chromosome separation mechanisms by unlike organisms. Some of these may take been intermediate stages in the evolution of the mitotic spindle of college organisms. For all examples except leaner, but the central nuclear region (more...)
Eucaryotic tubulin and procaryotic FtsZ clearly take a common evolutionary history. But, microtubules are important for chromosome segregation in even the most archaic eucaryotes, where they are also present in flagellar axonemes (discussed in Chapter 16). Whether the flagellum or the spindle evolved first is unclear.
A somewhat more avant-garde, although still extranuclear, spindle is seen in hypermastigotes, in which the nuclear envelope once more remains intact throughout mitosis. These large protozoa from the guts of insects provide a particularly clear illustration of the independence of spindle elongation and the chromosome movements that separate the chromatids. The sister kinetochores get separated by the growth of the nuclear membrane (to which they are attached) before becoming attached to the spindle. Only when the kinetochores are near the poles of the spindle practise they acquire the kinetochore microtubules needed to attach them to the spindle. Because the spindle microtubules remain separated from the chromosomes past the nuclear envelope, the kinetochore microtubules, which are formed outside the nucleus, must somehow attach to the chromosomes through the nuclear membranes. After this attachment has occurred, the kinetochores are fatigued poleward in a conventional manner (see Effigy 18-41).
Organisms that form spindles inside an intact nucleus may represent a further stage in the development of mitotic mechanisms. In both yeasts and diatoms, the spindle is fastened to chromosomes by their kinetochores, and the chromosomes are segregated in a way loosely similar to that described for creature cells—except that the entire process generally occurs inside the confines of the nuclear envelope (come across Figure 18-41). Information technology is thought that the "open" mitosis of higher organisms and the "closed" mitosis of yeasts and diatoms evolved separately from a mutual ancestor resembling the modern hypermastigote spindle. At present, there is no convincing explanation for why higher plants and animals accept evolved a mitotic apparatus that requires the controlled and reversible dissolution of the nuclear envelope.
Summary
Cell division ends as the cytoplasm divides into two by the procedure of cytokinesis. Except for plants, cytokinesis in eucaryotic cells is mediated past a contractile ring, which is composed of actin and myosin filaments and a diversity of other proteins. By an unknown mechanism, the mitotic spindle determines when and where the contractile ring assembles and, thereby, when and where the cell divides. Virtually cells split up symmetrically to produce two cells of the same content and size. Some cells, however, specifically position their spindle to divide asymmetrically, producing two daughter cells that differ in size, content, or both. Cytokinesis occurs by a special mechanism in higher-constitute cells—in which the cytoplasm is partitioned by the construction of a new cell wall, the cell plate, inside the cell. The position of the cell plate is adamant by the position of a preprophase band of microtubules and actin filaments. The organization of mitosis in fungi and some protozoa differs from that in animals and plants, suggesting how the circuitous procedure of eucaryotic cell partition may accept evolved.
Source: https://www.ncbi.nlm.nih.gov/books/NBK26831/
{kind=link}
Post a Comment for "The New Cells Must Immediately Begin to Divide Again"